

THE TAKE AWAY: Does insulin and melatonin work by taking its cues from solar spectrum? Yes, they do.
If so, how is insulin’s action quantized?
When energies of electrons are higher because of the action of the photons of the sun, that signal is transduced by the cytochromes in the electron chain transporters. Most electrons from carbohydrates enter at cytochrome one because NADH is a fluorophore-protein. It absorbs best at 340 nm range of light which is strongly in the UV range of the solar spectrum. This couples photoelectrically with insulin release from the pancreas. It uses another solar hormone, melatonin, to help it achieve its goals in the gut. Melatonin is first made with the interaction of UV light and aromatic amino acids in the eye. I covered how melatonin links to the sun and to time in Time 9.
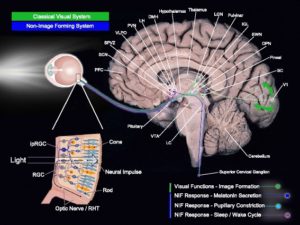
Figure 1.Fig. 1
Light-sensitive brain pathways. Simplified brain networks (not exhaustive representation) of the classical visual system and the non-image-forming system. Abbreviations: PFC prefrontal cortex, SCN suprachiasmatic nucleus, SPVZ subparaventricular zone, VLPO ventrolateral preoptic nucleus, PVN paraventricular nucleus of the hypothalamus, LH lateral hypothalamus, DMH dorsomedial nucleus of the hypothalamus, LGN lateral geniculate nucleus, IGL intergeniculate leaflet, EWN Edinger-Westphal nucleus, OPN olivary pretectal nucleus, SC superior colliculus, V1 primary visual area, LC locus coeruleus, VTA ventral tegmental area, ipRGC intrinsically photosensitive retinal ganglion cell, RHT retino-hypothalamic tract.
Melatonin is produced first by the eye and then ocular melatonin helps liberate melatonin in the pineal gland in response to the absence of light. If light, especially from 400nm -465nm is present none is released in the pineal. Melatonin levels are quite low during the day and reach their peak 4 hours after darkness. Like other hormones, melatonin works by receptor-mediated bonding using calcium as a controller. People like to think its a type of a lock and key mechanism but this is factually incorrect. It is more like an electric gate control we see in semiconducting light diodes. Melatonin receptors are found to a great degree in the retina, RPE, and the visual pathways of the brain. When light is absent then melatonin activates this “light diode like” gates.

For humans, the absence of light is an optical nonlinear signal that indicates that it is the period when redox levels are unstressed by solar radiation. This is how the brain tells time, night from day. It is an old evolutionary clock that was important before the SCN came along. This clock, however, in humans still controls glucose metabolism. It also is the reason why blue light at night is what is driving insulin resistance and the runaway diabetes we see today. The optical signal of melatonin is a key circadian timing signal contributes to and is a part of a cascade of other responses that help initiate and maintain sleep when light is not present. Melatonin levels also control the distances between respiratory proteins in our mitochondria. If the level is low anywhere in the system % heteroplasmy rises and the ATPase of that mitochondria begins to spin slower than it should and electron chain transport slows in that tissue and damage occurs that can make that organ diseases.
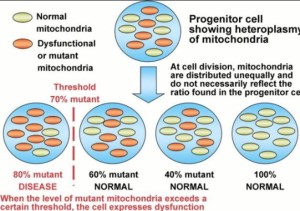
What is mitochondrial heteroplasmy?
A large number of pathogenic mtDNA mutations have been identified and the more severe mutations are frequently mixed with normal mtDNAs within the cell, a state known as heteroplasmy. Heteroplasmic alleles can shift in percentage during both mitotic and meiotic cell division, leading to a potentially continuous array of bioenergetic defects, a process known as replicative segregation. As the percentage of mutant mtDNAs increases, the resulting bioenergetic defect becomes increasingly severe. Because different tissues have different bioenergetic thresholds, as a patient’s bioenergetic capacity declines it eventually falls below the minimum threshold for that tissue and symptoms ensue. Because the tissues and organs with the highest bioenergetic requirements are also those that are primarily affected in the common metabolic and degenerative diseases, it follows that mitochondrial dysfunction may be a major contributor to complex diseases.
Women that harbor deleterious heteroplasmic mutations have a high probability of having affected children, the nature, and severity of the phenotype depending on the mtDNA mutation and the percentage of heteroplasmy. Cells and individuals can accumulate an array of different mtDNA mutations over time, the aggregate of which degrade the energetic capacity of the cell. Such mutations are important in aging and cancer. Given the enormous potential explanatory power of heteroplasmic mtDNA mutations, it is striking that very little is known about the origin, genetics, and phenotypic effects of heteroplasmic mtDNA mutations.
Strangely enough, a researcher now knows that MELATONIN receptors are also found in the pancreas, but clinicians have no idea this is what is behind the diabetes epidemic globally. Melatonin and insulin receptor density are brisk in pancreatic beta cells. This is how diabetes manifests in quantum fashion from environmental exposure of light at night via our eyes or skin. Most people know beta cells release insulin during the day when we are supposed to eat. This points out why My Leptin Rx tells you not to each once the sun sets. If you do you ruin the circadian control of the melatonin cycle. Beta cells regulate glucose levels in the blood. The LCHF folks focus on insulin alone and completely forget about melatonin and the effect of light at night on glucose metabolism.

During the daylight blood pools to the surface to absorb specific frequencies of sunlight while releasing nitric oxide. Researchers also know that when melatonin activates these receptors via the blood plasma, insulin secretion is decreased by the pancreas. This mechanism shows you diabetes is a photo-electric disease related to protons and electrons.
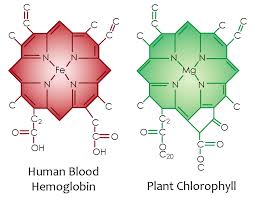
Here you can see the atomic lattice of nitrogen around different metals. Nitrogen electrons are excited by sunlight and this energy is used by the metal inside to create electric currents from sunlight frequencies.
Photoelectrically you say? In plants, sunlight hits a magnesium atom at the center of chlorophyll to liberate an electron that is immediately ferried away to a reaction center called P680 to liberate electrons from water. The first step in photosynthesis is the oxidation or BURNING of water to liberate electrons. When wood burns in air, oxygen atoms pull electrons from carbon atoms. The electrons in carbon are lovely attached so carbon burns easily. When a plant or animal burns water the process requires a lot of energy. all photosynthetic reactions are unique in that it is the only place in the cosmos where water is burnt to yield electrons. So where do these electrons go in plants? NADPH. This protein is an electron and proton carrier in a cell. Where do they go in animals? NADH and NADPH. They both carry electrons and protons in animals. That is the redox couple in cytochrome 1 of mitochondria. These electrons are ferried to oxygen as the terminal electron acceptor in animals. The protons recycle from the matrix of mitochondria. In this quantum dance, protons, are moved by red light, water is charge separated by UV and IR light, and ATP is made in several ways. One from a protons force within the mitochondria and another from the penetrating ability of red light in sunlight. If ATP is not made abundantly insulin resistance is the result. Very few people seem to realize that a lack of full-spectrum sunlight can drive insulin resistance. They think insulin resistance is a carbohydrate story. It is not. Light is the most powerful effect on insulin and melatonin. When these signals are corrupted your time on Earth is shortened by some disease tied to the % heteroplasmy in your tissues somewhere. Do diabetics with metabolic syndrome live longer or shorter than those without the mitochondrial condition? Insulin resistance is not a food only problem, it is a light at night problem.

Tyrosine is critical to insulin’s release and its storage. Tyrosine is an aromatic amino acid that absorbs UV light by nature’s design. I bet you have not heard a food guru tell you this before. They have not considered the quantum mechanics of life. The energy is given off to these mitochondrial proteins and this causes specific phosphorylation of these proteins to alter their ability to function. NADH/NAD+ are the redox couple buried in cytochrome 1.

This means that the highly energized spring/summertime electron signal is coded for by the hormone insulin. When these types of electrons are sensed in the blood plasma, insulin is made by the pancreas. These electrons get excited by the solar radiation during the day. When insulin is made, we see electrons being shuttled to complex 1 of the ETC, with a resultant large supply of NADH also being made at complex 1. If UV light is not present within a cell when these electrons are added to mitochondria NAD+ begins to lower. This slows ECT tunneling of electrons. When enough UV light is present in our cells and carbohydrates are broken down to their electron form things work properly. When this occurs, we also have a restricted supply of fatty acids being delivered to our mitochondria from our fat cells because of the action of insulin on adipocytes. Insulin only has these controls when carbs are eaten and ELF-UV is able to be properly released from a cell. When this happens, it generates a high membrane voltage in our inner and outer mitochondrial membrane. This voltage is a charged energy that the electromagnetic force controls. This is the only force in nature that controls charged subatomic particles like electrons and protons. It turns out cytochromes use protons too.
If you watched any electromagnetic force video on youtube, you’ll see that the electromagnetic force controls particles of atoms when they carry a charge. Protons have a positive charge and electrons have a negative charge. These are the particles that the force acts upon us in our mitochondria.
Many scientists in the 1950’s were 100% pre-occupied with finding out where the high membrane voltages came for in life because he knew they were quite important for many processes. One, named Gilbert Ling, was the first to find out that it was not due to the membrane pumps. Today, biochemistry still thinks these pumps are the source of the 30 million volt charge in mitochondria. Photosynthesis has shown us where it really resides. It is a function of Lady Evolution’s semiconductor design in chlorophyll. Life’s battery is all about the charge separation of water into electrons and protons. That is the first step in photosynthesis and the it is first step in animals cytoplasm around mitochondria too. The mechanism is different but the battery it creates explains the energy deficits that membrane pumps cannot explain mathematically. How so? It uses water as it medium to a sea of free electrons and controls the flow of protons in cell water.
Dr. Doug Wallace talks about this in his experiments when he put healthy and damaged mitochondria with healthy and sick cell cytoplasms. The result was that the cytoplasm drove illness and the cytoplasm responds to the environment and the exclusion zone (EZ) memorized its signals. EZ water is a sea of coherent free electrons that can store energy and information in protons. Pollack’s work on EZ is well known now by bio-physicists but not by clinicians. Del Giudice and colleagues pointed out 50 years ago, most if not all water, in living organisms is interfacial water. This means it is almost never further away from hydrophilic surfaces such as membranes or macromolecules than a fraction of a micron. Gerald Pollack went further with this work to describe the physical characteristics of water in the EZ experiments. EZ water = coherent water. Because coherent water is excited water with a solar plasma of almost free electrons, it can easily transfer electrons to molecules on its surface or mitochondria which swim within this sea of EZ. Those signals are optically transferred over EZ hydrogen bonding network to the MINOS which controls the respiratory proteins size and shape. How? Del Giudice and colleagues also published that water coherent domains (CDs) can be easily excited by light, and are able to collect small external excitations to produce single coherent vortices whose energy is the sum of all the small excitation energies, turning the originally high entropy energy into low entropy coherent energy, which is trapped stably in the water CDs. This coherent energy in turn enables selective coherent energy transfer to take place as follows. It turns out melatonin controls this process optically at night. All molecules have their own spectrum of vibrational frequencies. Everything protein in life is a vibration, a color, a frequency. If the molecule’s spectrum contain a frequency matching that of the water CD, it would get attracted to the CD, and become a guest participant in the CD’s coherent oscillation, and settle on the CD’s surface. Furthermore, the CD’s excitation energy would become available to the guest molecules as activation energy for chemical reactions to take place. Melatonin has this very effect at night with respect to water. This is why most dehydrated people will have altered melatonin cycles. Clinicians can see this on their BUN/creatinine ratios if they bother to look. This selectivity may be the reason why out of a hundred different amino acids only 20 have been selected for making proteins in living organisms. This frequency signal in the cytoplasm determines the free radical signals made and the ultimate response of energy flows to and from mitochondria.

The membrane pump theory breaks the second law of thermodynamics by 500 fold when we add up how much ATP a cell can make versus how much it uses. This means we have to make energy from some other place. Water, sunlight, and protons make up the big difference. ATP is important but it is not the key to a cell’s bio-energics.
This voltage inside our membranes is what the electromagnetic force pays deep attention to, as I mentioned to you in Energy and Epigenetics 6 blog. This voltage is directly linked to the cell’s ultimate redox power and redox power is linked to how much light is collected in EZ water in a cell. When the voltages are high in membranes anywhere, but especially in the mitochondria, the result is a “relatively stronger current of electrons down the ETC via complex 1. Because the current is strong in one only direction during summer, no reverse electron flow can happen at complex 1, because there is a minimal electron current simultaneously occurring via electron transporting flavoprotein dehydrogenase’s FADH2 at complex 2. This is where fat electrons enter ECT and where protons are stripped of food. This excess energy from summer time foods is stored as fat when UV light is absent from our environment. This is why carbs can fatten all animals in autumn to ready them for winter. Humans face this too. It is not carbs that fatten us, it is the lack of UV light that is the photoelectric signal that does. This is how we store excess electrons, or highly energized electrons, which are elevated in their energies by the photoelectric effect of the summer light’s power in fat stores. When the electromagnetic force of light is stronger in your environment from non-native solar sources (blue light from tech gadgets) , the results are they we gain more weight because we slow ECT transport. We uncouple insulin action from UV light exposure. Non-native EMF causes us to make excessive NADH in our mitochondria causing us to crave carbohydrates. If we eat them when no UV is present, we lower NAD+, create pseudohypoxia because ECT slows. All electrons are delivered to oxygen to reduce it. The only reason we breath oxygen is to capture electrons. Not too many people realize this.

After the photoelectronic signal for insulin release has been produced in a cell, termination of signaling is also then needed by a cell. How does this happen? Insulin is degraded by endocytosis and degradation of the receptor bound to insulin as a main mechanism to end signaling. It turns out at the center point of insulin molecular structure is an atom of zinc, a transition metal, surrounded by 4 specific amino acid residues with amide linkages. These residues all are quantized to specific light frequencies and are affected by an aqueous surrounding. If those light frequencies are not present insulin never gets degraded properly. The human insulin protein is composed of 51 amino acids and has a molecular mass of 5808 Daltons. It is a dimer of an A-chain and a B-chain, which are linked together by disulfide bonds. Within vertebrates, the amino acid sequence of insulin is strongly conserved. Protons and electrons can change how it operates. In most species, the A chain consists of 21 amino acids and the B chain of 30 amino acids. Insulin molecules have a tendency to form dimers in solution due to hydrogen-bonding between the C-termini of B chains. Additionally, in the presence of zinc ions, insulin dimers associate into hexamers. These interactions have important clinical ramifications. Monomers and dimers readily diffuse into blood, whereas hexamers diffuse poorly. Sunlight affects these conformational changes. Protons motions are affected the biogenic amine amino acids. Insulin has many of these amino acids in its beta chain. There are five established biogenic amine neurotransmitters: the three catecholamines—dopamine, norepinephrine (noradrenaline), and epinephrine (adrenaline)—and histamine and serotonin. Most of the biogenic amines are made from aromatic amino acids.
This, in turn, affects the proton signature at the amide sites on the amino acids. Its release stays in the constant on position until the beta cells of pancreas burn out photoelectrically because no stem cells are left to replace them. The interaction of the electromagnetic signals we allow from our environment and the interaction of zinc and these specific amino acid residues causes a change in the electronic signal to induce dephosphorylation in insulin. This removes energy from electrons or it removes electrons from these proteins to induce dephospho rylation. Blue light keeps the switch for insulin release on. UV and IR light terminate them on the switch. What happens if you are deficient in UV and IR light in your environment and get too much blue light at night? You get a form of diabetes.
IS THERE ANY ANALOGIES IN SCIENCE TO SHOW US THIS MECHANISM?
If you watch the electromagnetic force videos on you-tube you will see semiconductor engineers add phosphate to silicon wafers to increase the sheer numbers of electrons to the silicon backbone to pre-load it for more optimal functioning. This makes a “N type” semiconductor. An N type semiconductor is negative because the added electrons from phosphate have a negative charge. The more phosphate you pre-load a semiconductor with, the more negative charge you get. In our excitable cells, our back bone of our semiconductors are not made of silicon, but of carbon surrounded by water. Being a bit more specific, our backbone is the triple helix of collagen surrounded by an ocean of intracellular water in our quantized cells to generate life. Water also happens to be the “N type” semiconductor. It collects electrons from all sources and distributes it throughout our cells and organelles. Health is a function of how many electrons your semiconductors have. Electrons are the only way UV light can interact with our proteins photoelectrically. No other frequency of light is capable of doing this. When electrons are added to proteins or lipids they become more hydrophilic. They love water more than normal. This is important because the charge separation is the main source of power inside a cell.
When water is adjacent to collagen (hydrophilic substance), it separates into negative and positive charged atoms of protons and electrons. This is the what my Energy and Epigenetics 10 blog showed you in picture and word form. Water has a fourth phase when it is placed adjacent to a hydrophilic substance. It forms a negatively charged exclusion zone. This is how the battery inside you cell first forms. After water touches the hydrophilic proteins a small EZ is made. IR light makes it grow. UV light makes it massive. It has been experimentally proven by many scientists that this occurs in water and in the light. Most notable of them all is a Dr. Gerald Pollack. He decided to study this because of Gilbert Ling’s work in the 1950’s on water and insulin. This charge separation creates a negative and positive semiconductor to develop within EZ water and collagen suspended within this battery acts like a wire that connect both oppositely charged semiconductors in water to form an electric circuit. If you carefully watch the video on the electromagnetic force, you will see this emerge in your own mind. What forms the semiconductors in you is very counter intuitive if you are a biologist. If you have no such educational biases you can see precisely how water works inside a quantum cell. But those things are the three fundamental things in Nature I spoke about in detail in Energy and Epigenetics 4 blog.

How does a cell get rid of insulin’s actions in a quantized fashion?
The signaling of insulin is terminated by dephosphorylation of these tyrosine residues inside of insulin’s molecular structure. Tyrosine is an aromatic amino acid that absorbs all forms of UV light and can be programmed by protons in the blood plasma. The process of dephosphorylation is tied to the flow of electrons on the tyrosine residues by tyrosine phosphatases. We have known for some time that serine/threonine kinases will also reduce the activity of insulin, but few people have realized how the control of electrons and protons dictates the mechanism of action. Proton motions are affected by temperature changes. Ling found insulin stops working optimally at 62 degrees F. This is likely mediated by protons tunneling transfers from the blood plasma irradiated by the sun in the skin. There is a quantum another effect on these electrons by the light in our environment.
It turns out that temperatures have the exact same effect on these residues by inducing the Hall effect on electrons. Heating favors H+ proton transfers. Cooler temperatures inactivate the effect of insulin and this changes the solar spectrum of light too. This is why cooling often transiently raises our blood sugar. Some animals outside of the tropics use the glucose as an antifreeze for their blood to help them get through winter when it freezes. Their bodies do, but their blood does not. This helps explain why Gilbert Ling found in 1969 that insulin action on D-glucose varies inside of cells when the temperatures falls below 62 degrees F.
The final action of insulin termination is associated with the reduction of the number of receptors on the plasma membrane. When coolness is present or there is photo-electronic induction is altered by light frequencies, there is a decrease in the amount of insulin receptors present in our cell membranes which also brings to a termination of insulin signaling. This defines physiochemical thermoplastic control of a biologic response I spoke about long ago in the Cold Thermogenesis 6 blog post. This is an example of how the same molecular key can open both the door to heaven and the gates to hell depending upon the context in which the “key” is found.
KEY POINT: If you are following this post well, we would expect to see higher levels of NADH being delivered and developed in our mitochondria when non-native EMF is present to an excess. This is why modern technology gadgets using nnEMF frequencies are not good for those with diabetes. There use would simulate a constant season of “summertime electrons” to our mitochondrial proteins 24/7. This would act to slow ECT while depleting our beta cells of all their insulin.

When we starve, we use our own fat stores to make ketones, which act to drop NADH levels, raise NAD+ levels, because we are using protons liberated from fat stores. We are also feeding electrons from those fat stores into FADH2 and electrons at cytochrome 2. So this simply explains why non-native EMFs make you think you need carbs, so the vagus nerve creates a carbohydrate craving response in your brain by raising a chemical called neuropeptide Y. These chronic actions also stimulate mitochondria that can no longer make superoxide well. We call this heteroplasmy. We can handle some heteroplasmy but as its percentage rises there becomes a critical mass where a mitochondria must be replaced because ECT is too slow. We need a superoxide burst to make this signal. In diabetics, this signal is lost. The reason for this is complex quantum signaling in the matrix with electrons, protons, and oxygen levels. Their tissues are forced to work with defective mitochondria and ECT remains forever slowed. When you do not understand this quantum dance you believe that foods, specifically carbohydrates cause diabetes when the reality is that they just amplify the signals in bad mitochondria. Slow ECT is what is behind most metabolic diseases and diseases of aging. The percentage of heteroplasmy in a cell is far important than the foods we eat. Lowering our carbohydrate burden helps but we have to add back UV light from the sun because this can help us regain our superoxide burst.
This should explain to you what I wrote in the EMF 4 blog post now. Non native EMF elevates our perception we need carbs because it raises our heteroplasmy percentage dramatically by dehydrating cells and cause loss of calcium in our mitochondria. This causes mitochondria to swell and slows electron and proton tunneling even more. It worsens the % of heteroplasmy, and melatonin and insulin signaling do not operate as they should. This is why the light environment we allow is more critical than the foods we eat. We never get this message because no one is educated well about the quantum actions of light in proteins or mitochondria. It also explains why and how uncoupling proteins work as they do in mitochondria. They are examples of photonic “Maxwell demons” who pay deep attention to the energies in the electrons and photons delivered to the inner mitochondrial membranes via water networks mentioned earlier. When energy is high we should be able to uncouple to get rid of excess electrons or their energies as free heat. This heat shrinks the water around mitochondria while shrinking the respiratory proteins and improving tunneling speeds of both subatomic particles. This is why statins can cause diabetes. Statins block ECT tunneling from cytochrome 1,3, and 4 because it blocks the Q-cycle in mitochondria by disrupting CoEnzyme Q10 function. This indirectly affects proton tunneling. So uncoupling naturally increases when our mitochondrial membranes sense a higher voltage for any reason. This is why we burn free heat (IR light) when we are healthy. When this occurs your inner mitochondrial membrane will oscillate at 100Hz. Your muscles mitochondria will too. Then and only then can you burn fat by beta oxidation. Fat burning has ZERO to do with your fuel choice, it has 100% to do with how your mitochondria vibrates. That is controlled by the battery stored in the EZ of water around a mitochondria and that battery is linked to environmental light signals. Because of this quantum relationship, everyone has a DIFFERENT efficiency in pumping protons out and converting them to ATP—this is called the coupling efficiency!! It is related to your mitochondrial haplotype and your location on Earth. Today it is massively effected by nnEMF due to population density. Before nnEMF, people that are very efficient at pumping protons out will require the MINIMUM amount of calories to create the Maximum amount of ATP. So every time you burn a calorie you will generate a MINIMUM amount of Heat for a MAXIMUM amount of work. An inefficient proton pumper will require MORE calories to pump the SAME number of protons and convert them to ATP. Since you’ve used MORE calories you create MORE heat. The more heat produced the more leaking of IR light occurs around mitochondria to give us 100Hz frequency. The heat improves the motions and movements of H+. This improves the cycling in the TCA and urea cycle. Today, our modern world has trashed these natural relationships.

This IR heat shrinks the water around mitochondria to increase ECT speeds, improves the spin rate of the ATPase and cycling in the TCA and Kreb’s cycle while lowering the % heteroplasmy in the mitochondria. That is the quantum jazz in your cell. It is also why our core temperatures are so linked with how well or ill we are clinically. Cooling our surfaces causes us to release more heat from our mitochondria to improve their function by getting rid of bad ones. This is how heteroplasmy is handled in winter. The story is a bit more complex with uncoupling proteins because we can actually use insulin to uncouple in our species too!
The electromagnetic force present in light also turns on and off melatonin via the action of electrons contained in melanopsin in our eye using a similar manner. I covered those details in Ubiquitination 24 blog if youre interested. It is also really teased out in my current Time series. When ocular melatonin is made, it couples with adenosine release in the brain to induce sleep. Ocular melatonin also needs UV light stimulus during day time and complete absence of blue light at night to work. It is only then that the serotonin stores in the small bowel can be harvested to make make pineal melatonin from the gut stores of serotonin. The stimulus of this complex program is UV and IR light during the day via the RPE of the eye, with no blue at night, while the there is no peristalsis in the gut. This is why eating late at night in blue light destroys our sleep and makes us fat. It also happens to keep our insulin quantum switch constantly on. It is not insulin that makes us fat. It is all these uncoupled cycles occurring at once that do by ruining signaling. Sleep is 100% a light story. Quantum sleep is induced as I posted in Energy and Epigenetics 9. It is made most efficient when we are cold, ketotic, reduced because we are filled with electrons that initially charge separate water in our cells. The cell becomes reduced by making large amounts of glutathione when melatonin is high in the absence of the electromagnetic force of light. Glutathione is the main chemical that provides for all redox reactions in the body. Glutathione is also quantized to UV light by having the ability to quench redox reactions. I covered this in Time 6. This means the redox potential is 100% tied to your ability to live a long life healthy. It also means the sun is your friend………Vitamin D3 and insulin have a lot more in common than a surface understanding gives you. Biochemistry only provides substrate understanding, but biophysics controls how they work. Light brings its true focus into view. Insulin release is 100% quantized and modern biochemistry does not yet fully understand this. You must.

CITES
Included in hyperlinks.