

THE TAKE HOME: Can light release from a cell tell us about mitochondrial energy status? Is mitochondrial fission involved in the elimination of ROS-producing mitochondria? Can our cellular light fingerprint tell us about this process I call “mitoptosis”? Mitochondria self-regulates their function by participating in the multi-faceted defense of the cell against oxidative damage. When the cell is under siege by ROS/RNS it releases ELF-UV and this light is the signal for the recycle programs in mitochondria designed to buy us more time on earth.
HOW MUCH DO YOU KNOW ABOUT CELLULAR LIGHT RELEASE?
When one reads through the Roeland van Wijk’s book one could easily conclude that cancer is more a mitochondrial metabolic disease in accordance with the original theory of Otto Warburg. Today’s war on cancer was waged in 1971 by Nixon and this war has been waged on nuclear DNA changes. So far we have no cure and we have spent 1.3 trillion dollars since that time looking under the wrong genome. In the cytosol, there is another genome in humans. That genome is 37 genes large and codes almost exclusively for the respiratory chains in electron chain transport. Today modern oncology continues to cling to the belief that oncogenesis is the product of point or accumulated somatic mutations. The recent work (forty years) of Dr. Doug Wallace seriously blows this viewpoint up. VIDEO
His video 50:00-59:00 really makes the key points for modern humans. All diseases are linked to the geometry of the respiratory proteins. As they spread apart new disease phenotypes emerge from the change in distance without ANY NUCLEAR GENOMIC changes.
This viewpoint is 180 degrees to all paradigms in modern medicine.
Is this why we can’t solve the puzzle of neolithic diseases? Are we looking in the wrong place for the problem? Today’s paradigm is built around the old ideas of anatomy and Mendelian genes and not where energy is generated. Might our bio-medical premises of the 20th century be outdated, and frankly, incorrect? Physicians are still taught to think about the disease from the anatomical perspective that tissue level disease contain anatomical defects. When tissue defects cannot explain the problem we jump straight to 1953 when DNA was discovered and we blame defective genes. We’ve been doing this for 70 years and it has gotten us sicker and in more debt. Physicians are not taught to look at energy defects as the cause of tissue level disease. This must change.

The belief began with a simple half-truth: All genes are chromosomal, therefore, we must have Mendelian defects present at some level so we look into the nucleus of the cell for the answer next. Mendelian genetics is only 125 years old. What if both ideas are terribly flawed because they both ignore the key concept in life, namely energy generation? In a cell, energy is transformed into the cytosol allowing life to manifest. Beta oxidation makes exponentially more energy in the cytosol and the Kreb’s cycle can in the mitochondrial matrix. The more your mitochondria can fat burn the less disease you’ll get. Today doctors and patients think fat burning is due to the fuel choice we eat. It turns out this is 100% wrong too. Fat burning is 100% linked to how the inner mitochondrial oscillates based upon the light that collides with it and released from it. This idea is contrary to medicine’s core tenets. It is very counterintuitive to the foundation associations that have become marketing machines for diseases. So if the disease never gets reversed, your paradigm is guaranteed a constant source of revenue.
In today’s medicine genetics has corrupted our ability to think. We’ve allowed genetic knowledge to narrows our perspective in healthcare, and we’ve forgotten that clinical experience broadens it, by raising your curiosity. Since modern healthcare cannot solve chronic disease, you’d think they’d be interested why they keep swinging and missing. They are not. Just look at how we fund research. No one wants to talk about how nnEMF and blue light alter the programming of electrons in proteins because our economy now is built on it.

These ideas are very disruptive to the cancer machine created by organizations. Cancer is now a huge business in healthcare systems. The ‘Warburg effect’ was an observation made long ago that cancer cells produce energy by a high rate of glycolysis followed by lactic acid fermentation in the cytosol. The cytosol is where the mitochondria lie not the nucleus. This process is also associated with massive loss of ELF-UV light from cells. That data came after Warburg’s biochemical observations. In normal cells, there is generally a low rate of glycolysis followed by oxidation of pyruvate in mitochondria. Eukaryotic cells are designed to use beta-oxidation as their key pathways to delivering energy/information to tissues to maintain their tissues anatomy or organization.
LIFE = STRUCTURE + ENERGY + INFORMATION
All things in chemistry are linked to what happens to the valence electron in an atom. It turns out all things in life are linked to what happens to valence electrons in what matter makes life up. This is how the photoelectric effect works in plants and animals.
Is there a trend in the Periodic Table for threshold frequency in photoelectric effect that life takes advantage of?
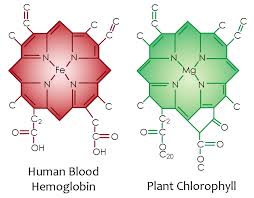
There are trends for reactivity in the periodic table, so there is an obvious trend for the threshold frequency of the elements. This trend teaches us about life’s choices in proteins. Does this trend have any bearing on the threshold frequency of compounds formed by these elements in animals and plants? Look at the chemical compound of chlorophyll used for photosynthesis in plants and hemoglobin in the animal blood. They are almost identical with the only difference the central metal atom. How does this link to the photoelectric effect?
The “threshold frequency” is commonly known as the work function, symbolized by F in Einstein’s equation for the photoelectric effect:
Eke = hn – F
F represents the energy required to knock an electron lose, and we would, therefore, expect any trend to correspond to electronegativities or ionization energies. Remember that E=hn, so that the lower the work function, the lower the frequency of light required to knock an electron lose. This is why plants use magnesium (Mg) in chlorophyll and why humans have iron (Fe) in their hemoglobin. Mg has a work function of 3.66 eV and Fe range hovers around 4.67-4.81 eV. This means the blue light is capable of knocking electrons out of Mg atom to make an exciton with ease but it cannot do it to heme proteins with iron. Recall that plants evolved before animals by about 50 million years. Also recall that not all frequencies of sunlight are present during all light hours with the exception or Red, Green, and Blue. Blue frequencies are always balanced by these other frequencies and this is why the photoreceptors of the camera vision of the eye only use these three colors to all us to see. So why did plants decided to use magnesium chlorophyll? Blue light from the sun is always present at all hours of the day and since trees and plants cannot move across the tectonic plates; so it makes sense their photosynthetic energy systems are based around magnesium chlorophyll and not iron chlorophylls. It also points out why humans need strong UV light every day at some level to keep autophagy and apoptosis working to self-regulate themselves. UVA light is present at most hours the sun is shining. UVB light is more restrictive when it is present.

This is why animals and their eyes and skin have evolved a retinal pigmentum epithelium in the eye and melanin on their skin. This chemical is the photosynthetic pigment that absorbs all UV frequencies.
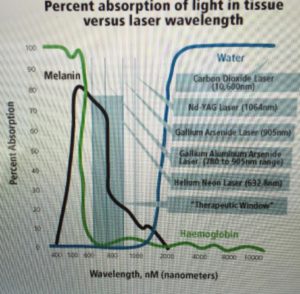
Recall UVA/B light frequencies is not always present at all times of the day and since humans can move they can access stronger light to capture more energy in their pigmented blood proteins and mitochondria. This is also why melanin is a biogenic amine found in animals skins and eyes. The more UV light they get the more photoactive melanin becomes. Melanin is the mammalian photosynthetic chemical. HYPERLINK
Those things that interact with sunlight photoelectrically and this is why DNA and RNA only code for specific proteins. Their photo-optical abilities are what nature really cares about. They only work with sunlight when they are hydrated by an exclusion zone of water. All living things have electrons and all electrons everywhere in the universe are identical with one exception. Light excites them to give them different quantum numbers. Life is carbon-based. The quantum numbers used to refer to the outermost valence electrons of the Carbon (C) atom, which are located in the 2p atomic orbital, are; n = 2 (2nd electron shell), ℓ = 1 (p orbital subshell), mℓ = 1, 0 or −1, ms = ½ (parallel spins). Once light programming occurs, no electron is identical to any other in the universe. Before light programming, all electrons are the same everywhere in the universe. This means abiotic atoms are not programmed by sunlight and biotic electrons are. Your mitochondria were built by nature to decipher the information and energy in each electron it processes to build the structure that life needs to live. Food electrons are programmed by the sun because they were programmed by it before you consumed it. Sunlight has a power density and barcode that varies as seasons vary via latitudes. When you understand this about electrons, you see how sunlight truly sculpts what life is and what life can become.

Information is buried in the signals that mitochondria create and use to direct the structure of cells and tissues. All ROS and RNS are free radical signals all born in mitochondria. People seem to forget this very fundamental tenet of biology. This implies that tissue level diseases can result from global energy deficits. It also implies that non-Mendelian genetics that is prevalent in mitochondrial DNA, and this energy production directly controls nuclear DNA expression via the epigenome.
This is a huge paradigm shift in medicine because our entire intellectual library in medicine is being attacked by this new data from bio-energetics of mitochondria. Dr’s. Nick Lane and Doug Wallace are leading this charge on both sides of the Atlantic while most clinicians do not even know who these guys are. Our current constructs in medicine only reflect what we do believe to be true, but it lacks the insights to even ask the question what is it, that we do not know, or what we don’t know that we don’t know, that might be behind modern diseases? This question is rarely entertained because we focus solely on beliefs we think are true and mighty. What if all that we believe is based on a house of cards?
This is an unsettling idea to most, but when you understand that a neuro-hormone like melatonin controls mitochondrial DNA, you begin to see how melatonin controls the geometry of the respiratory proteins that control the flow of energy/information from a mitochondrial to run the cell. You begin to see why light and the absence of light at night is the most important thing for proper bio-energetics to occur in mitochondria.
If you understand the biology of light, the corollary to this concept is that light would then be seen to exert massive control over the expression of nuclear genes leading to many disease phenotypes. That has been what the basic science blueprint that has been published has foreshadowed for 100 years now. Modern corporate and government-funded data have avoided and ignored this inconvenient truth because of its economic implications. Our public problem is that our scientists and clinicians cannot fathom this just yet, and as a result, the public’s health is in a catastrophic state because our research and clinical focus is off the mark. This lack of focus is brought into sharp focus by modern mitochondrial science and biophysics. They have both been pointing to this inconvenient truth for 100 years, but the powers in biology, instead, have been mesmerized by the 1953 discovery of DNA and the ancient practice of anatomical dissection (Vesalius) to gain reductive insights about life and food macros. This is a bad way to view the modern world.

HOW DO LIGHT AND MITOCHONDRIAL SCIENCE MARRY TO ONE ANOTHER?
Conditions associated with pro-oxidant states, like cancer are states associated with elevated ROS production, demonstrate increased sensitivity to low-intensity light therapy (LILT). Cancer is very sensitive to low levels of light illumination. It is also well known that red light has massive effects on cytochrome 3 (cytochrome c oxidase) and ATP production.
Cellular sensitivity to red and infrared light occurs at the level of cytochrome C oxidase, is influenced by cellular redox state. Cellular growth phase, which also corresponds to cellular redox state, appears to be another determinant of this sensitivity. In vitro and in vivo, the effectiveness of LILT varies with cellular growth phase of cells. This makes light therapy in cancer very interesting if you look at it. Few do. Proliferating cells are in many cases more sensitive to incident light. HeLa cells (cultured cancer cells), fibroblasts, and epithelial cells all demonstrate sensitivity to LILT, which is more pronounced during the proliferative cellular growth phase of the cell cycle, and in each case, this proliferative phase is associated with elevated ROS production ant their oxygen utilization.
While there is nothing inherently wrong with the current thinking that a rapidly expanding list of mutated ‘driver’ genes and resultant brand of tumor they generate constitutes an explanation, something more is needed—something to unite and simplify. Since we have dry tapped cancer etiology for 50 years you would think people would begin to look into the cytosol where mitochondria exist. The idea that somatic mutations responsible for cancer spontaneously arise through random insult or error during DNA replication in normal noncancerous stem cells is certainly not implausible but based upon what we know from Wallace’s work it is highly unlikely.
LILT-induced improvements in traumatic brain injury (TBI) and cardiac conditions have been attributed by Oron et al. to rapid elevation of ATP content, increased angiogenesis. Making more RBC’s makes for more porphyrins and hemoglobin for light collection from the skin to offset the ELF-UV all human cells release to signal biochemical changes in cells. Porphyrins synthesis begins in the mitochondria. This is how light from the sun passes information on in cells. That information quanta only works for about 120 days in humans.
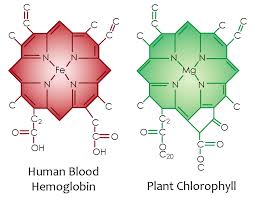
Here you can see the atomic lattice of nitrogen around different metals. Nitrogen electrons are excited by sunlight and this energy is used by the metal inside to create electric currents from sunlight frequencies.
Making more WBC’s allows for more melatonin protection for our mitochondrial DNA locally and globally within tissues. Sunlight stimulates this all if we allow it too. LILT also increases anti-apoptotic activity, heat shock protein formation, an increase of total antioxidants to stimulate regeneration cycles. Cancer detection has a quantum component that is being underutilized. A noninvasive redox measure that may be useful in advancing investigation of LILT in cancer and may one day be helpful in identifying responsive patients is the detection of elevated production and release of biophotons. It is currently believed biophotons come from ECT proteins and mitochondria. My belief is a bit different. The detection of biophotons, whose production is also associated with the production of ROS and the cellular redox state, represents just such an opportunity for clinical science. Over the last few decades, advances in photodetection have confirmed that many, if not all, living systems emit very low levels of visible and near-visible {ultraviolet (UV) and near-infrared (NIR)} photons that today are called biophotons. In cancer states, the amount of biophotons released is higher than in non-cancerous states. This light loss actually correlates perfectly with the Warburg metabolism he observed early in the 20th century. When ELF-UV light is lost cells must have intact control over apoptosis. Most do not have the ability to control autophagy. As a result, they become redox shifted.
Warburg’s conception of respiratory insufficiency (failure of the oxidative phosphorylation or OX/PHOS function of mitochondria) as the origin of cancer, its phenotypic descriptions, and even somatic mutation itself, not only has simplistic appeal, it can also be tested by today’s photomultipliers that measure light emission by cells.
There is another pathway of inquiry being used in metabolic and biochemical sciences.
Some of the primary evidence for Warburg’s theory now comes from nuclear cytoplasm transfer experiments in which so-called ‘cybrid’ composite cells are created in the lab. Cybrid experiments point to cell water as a huge clue in this process. These cells allow us to examine mito-nuclear coaptation. Tumorigenicity has been found to be suppressed when the cytoplasm of enucleated normal cells was fused with nucleated tumor cells. These experiments are a blow to the modern paradigm of cancer research that looks for cancer etiology within the nuclear genome.

ARE WE LOOKING IN THE WRONG PLACE?
In other words, normal cytoplasm, containing mitochondria from non-tumorigenic cells, are fully capable of suppressing the malignant phenotype of the tumor cell. This is where 99% of molecules are exclusion zone (EZ) water. Could it be a smaller EZ’s due to a lack of sunlight (UV/IR), are what cancers are fundamentally related too? It is well established by experiment that water is a repository for electromagnetic radiation to make a battery. It is also well established by Burr and Becker in plants and animals that regeneration is linked to the amount of a DC electric current. It has been well established by others that endogenous melatonin levels are present in people in the absence of tumor. We also know melatonin is made in the eye and skin via sunlight. This raises the interesting idea, might it be cancer comes from a lowered EZ and lowered DC electric current that turns off fat burning and forces a cell to use the more ancient glycolysis over the Krebs cycle to generate energies? Then it begins to make sense why glutamine catabolism is so unusual in cancer, doesn’t it? Is this why the urea cycle for glutamine is also increased? Why is the Kreb’s cycle in the matrix going unused in tumors yet electron transport upregulated? Doesn’t that situation create a massive spike in ROS which causes the cell to release a ton of ELF-UV? Is there a purpose to this set of circumstances we are not seeing? And about those two cycles, is something inside the cell blocking their shared use? Is the blockade due to something linked to the cytosol because of the Cybrid studies mentioned above? Cancer cells need brisk ECT transfer but they lose the ability to recycle via autophagy. It is almost like they have “lost their memory” for some reason. Could it be while energy is maintained information is being lost all at the same time? I think so because this is where the data leads me, not some “expert” opinion.
It appears the free radical signal, light frequency release, and electron/proton spin configuration of the free radicals are critical in this quantum dance. The amount of oxygen consumed is also linked to this reaction. The reverse situation of this experiment was also performed and is more problematic for the modern paradigm of cancer generation. When the “cybrid experiments” were reversed they showed that normal cell nuclei could not suppress tumorigenesis when placed inside a tumor cell cytoplasm and its tensegrity system.
Therefore normal nuclear gene expression, including presumptive normal tumor suppressor genes, doesn’t rescue malignant transformation. This is a big clue that where medicine is looking for cancer cures is clearly the under the wrong stone. The genome is not the problem but energy flows in mitochondria are. What links the matrix to the cytosol?
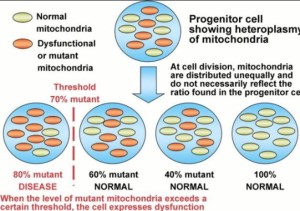
This may not necessarily prove that every road to oncogenesis has to pass through mitochondria any more than finding centrioles in dividing cells proves that every cell needs functioning centrioles to divide, but it does limit the scope of the somatic mutation conception. This should shift the research dollars to the mitochondria and biophoton research. So far to date, it has not.
A cytoplasmic origin of cancer, as opposed to a nuclear origin, does not mean that the problem has to be the mtDNA itself. It could be related to the lack of feedback control of the neuropsin/melatonin/DHA regeneration pathways. It can potentially be the result of a problem in the mitochondrial network that regulates everything from calcium efflux up to the scale of cell division itself. It might even be related to how water is created in the matrix.
We know that cell division requires the proper release of ELF-UV release at the precise time in the cell cycle and this is controlled by circadian clock genes that are linked to melatonin’s control of mitochondrial DNA. There is, however, some exciting new work that indicates disruption of essential OXPHOS proteins encoded by mtDNA induce metabolic reprogramming and cause the manifestation of the Warburg effect biochemically. In particular, it was shown that genetic silencing of cytochrome c oxidase by shRNA expression resulted in a metabolic shift to glycolysis, loss of anchorage-dependent growth in the tensegrity system of the cytosol, and acquisition of invasive phenotypes. It is also associated with a loss of the inner mitochondrial membrane oscillating at 100 Hz and this stops “effective beta-oxidation.” This suggests that defects in the mitochondrial electron transport chain can initiate a retrograde signaling cascade that leads to tumor progression. As this all occurs in a cell, when light emission is altered.

Photomultipliers have been used to further analyze biophoton emission from total organ brain and liver homogenates in cell fractionation studies These are not in vivo situations but they are instructive on light emission patterns. In these studies, mitochondria and submitochondrial particles have been shown to be predominant sources of cellular biophoton emission. This implies that increases in the distances between respiratory proteins lead to the higher emission of light from electrons on ECT or from protons inside cytochrome mouths that hit the iron-sulfur targets. We are not sure where the light is really coming from but we know it is from inside the cell.
Visible and NIR spectrum biophotons, linked to electron-excited states associated with the generation of ROS, are most relevant to LILT and to this blog. In contrast, the origins of UV biophotons are less clear today, and consequently, less clinically useful at this time. UV biophotons are currently theorized to be derived from DNA conformational changes (Popp/Kruse), amplified radical recombination reactions, delayed branch chain reactions in amino acids with aromatic amino acids, and/or direct fluorophore or chromophore emitters.
The data published to date suggest that singlet molecular oxygen is formed during free-radical processes in mitochondria. This often accompanies lipid peroxidation in cell membranes. This process is also controlled by eicosanoids under the control of the melatonin regeneration cycles mentioned in the Time 9 blog. Inside of cells, the outer mitochondrial membrane is a remnant of the endoplasmic reticulum of an early eukaryote. After 600 million years of evolution, this membrane is now connected to the endoplasmic reticulum in humans which is a membrane rich in DHA and eicosanoids. Both chemicals are known to be photoactive because of DHA’s massive array of electrons. This may be a major source of biophoton emission and why altered melatonin levels are associated with epithelial cancers and low sulfated Vitamin D3 levels. It points out a weak link in a cell that has cancer is a lowered UV environmental exposure. This limits electron chain speeds. This is not tolerated by cancer cells. It may be this is why cell has to re-emit ELF-UV to keep defective matrix level ECT operational because autophagy is defective. Interestingly, the ROS singlet oxygen is also thought to act as a signaling molecule in the cellular response to LILT photostimulation.

Biophoton emission has been related to the utilization of oxygen in many papers in the literature. These are covered in extreme detail in cite 1. So has the generation of ROS, and the production of electronically excited states in biological systems. Because of this, it has been suggested that biophoton detection could be used as a tool for the investigation of radical reactions and oxidative stress. Many biochemical techniques, which focus primarily on the measurement of biological lipid peroxidation, are now available to assess oxidative stress in living systems. Unfortunately, many of these techniques are quite invasive and inappropriate for clinical application. In contrast, biophoton emission provides, on a non-invasive basis, a signal of oxidative metabolism and ROS steady-state concentration that is readily and continuously detectable. It is possible to continuously monitor the metabolism of organs in vivo with biophoton detection techniques. We can do this with new camera’s today like the GDV camera Ruben and I used to develop the Quantlet.
Thus, in the assessment of in vivo redox status, biophoton detection may become more useful than other assays in cancer, including indirect assays of lipid peroxidation, such as glutathione release, electron-spin-resonance techniques, or assays assessing the evolution of hydrocarbons or malondialdehyde accumulation in tissues. These are all things we should be looking at in medicine using these new cameras but are not. Conventional wisdom and standards of care are barriers to progress. It is incumbent on the citizen scientist to begin to bio-hack these aspects of their biology themselves.
Visible and NIR biophotons and red and NIR LILT share some interesting parallels. Both involve visible and NIR photons, the mitochondrial electron transport chain, and the generation of ROS, in particular, singlet oxygen. As previously reviewed, LILT can stimulate cellular metabolism, presumably through photostimulation of elements (cytochrome 3) of the mitochondrial electron transport chain. Furthermore, LILT photostimulation is thought to result in electron-excited states and the increased generation of ROS.
Mitochondrial ROS generation is likely one of the primary mechanisms involved in LILT. As mentioned, Pal et al. demonstrated that, in human fibroblasts, low-intensity helium-neon laser irradiation-induced proliferation was associated with real-time transient increases in ROS production. Antioxidant mechanisms have been shown to inhibit such LILT effects, further implicating the role of ROS in LILT signal transduction.

The mitochondrial electron transport chain is a major source of visible and infrared biophotons. In fact, the metabolic processes involved in producing electronically excited states in biophoton-producing molecules are generally derived from oxidative metabolism accompanied by the production of ROS. Boveris et al. report that “hydroperoxide-supplemented cytochrome c provides a chemiluminescent model system suitable for the elucidation of some of the molecular mechanisms responsible for light emission”. Biophoton emission increases with increased generation of ROS by the mitochondrial electron transport chain.
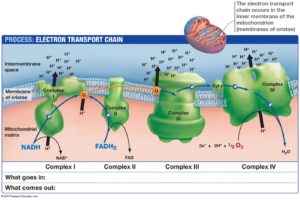
This light is then released down nanotubes filled with EZ water to transfer the energy and information by Fermat’s Law/Snell’s principal. Mitochondria have a superhighway of nanotubes that contain this light information for coherent transfer of the data to all parts of a cell and organism. Why is red light so special for mitochondria? The action spectra for an increase in DNA and RNA synthesis rate has been recorded when “cultured cells” are irradiated with radiation in the 400–860 nm region. The nucleus does not have chromophores absorbing in this region. Its histones and chromatin absorb in the strong UV range (350nm). They key is that these studies need to be done under direct sunlight outside of a lab to see the effect. Indoor light in a lab does not have these frequencies. This brings up the question why you need to question conventional dermatology and ophthalmology beliefs with respect to UV light being just a toxin and not a stimulant to nucleic acids in vivo. If UV light is so horrible for cells to ask your skin and eye doctors to explain how DNA has been shown in thousands of experiments to have a spectrum of fluorescence peak at 350 nm? Then ask them why the two proteins that are tightly associated with DNA, histones, and chromatin, both have been found by experiment to delay this UV light release in DNA in all normal cells? It is a fair question that none have answered for me.
Some researchers report that LILT and biophotons share another parallel. Like LILT, biophotons may influence cells through some form of photobiostimulation. At first glance, this is an appealing idea yet very difficult to conceptualize, as the two phenomena differ vastly in their functional intensities of light. Low-intensity lasers and LEDs are effective at an intensity window whose minimum is several orders of magnitude greater than the intensity of recorded biophoton emission which is in the ELF-UV range. In our current understanding, this makes it difficult to link biophoton-induced effects to LILT mechanisms because of the power and intensity differences. The key is, that light links both processes is some fundamental way. Light release in humans cells is oxygen linked. Data has confirmed that human biophoton emission is oxygen-dependent and reduced in hypoxic conditions. (Sinclair’s NAD+/pseudohypoxia 2013 paper)
What is the evidence you ask?
Just how specific are light frequencies on physiologic mitochondrial function?
The illumination of isolated rat liver mitochondria (nocturnal species) increased ATP synthesis and the consumption of O2. Irradiation with light at wavelengths of 415 nm, 602 nm, 632.8 nm, 650 and 725 nm enhanced ATP synthesis. Light at wavelengths of 477 and 554 nm did not influence the rate of this photochemical process. Oxygen consumption was activated by illuminating with light at 365 and 436 nm, but not at 313, 546 and 577 nm. Irradiation with light at 632.8 nm increased DWm and DpH, and as a result caused changes in mitochondrial optical properties, modified some NADH-linked dehydrogenase reactions and increased the rate of ADP ⁄ ATP exchange. Mitochondrial RNA and protein synthesis, replication of mtDNA as well as both cytosolic and mitochondrial protein syntheses were stimulated by He-Ne laser irradiation. He-Ne laser irradiation increased not only O2 uptake by cytochrome c oxidase but also electron transfer and proton pumping activity of this enzyme.
TENSEGRITY
Generally, in plants and budding yeast mitochondria are mostly positioned by actin filaments, while in higher eukaryotic cells and fission yeast they are distributed by microtubules. Neurons, do a little bit of both. To be properly inherited, fission yeast mitochondria must be reconfigured into a tubular network structure by microtubules. The intriguing thing here is that it now seems that yeast and mitochondria can move and be moved without the action of any motor proteins at all. We’ve known about this since 2003 as the cites show below. This is something we have suggested should be possible several times before, only now there is some evidence to show it. This idea stands in sharp contrast to the standard view which presumes that despite their comparatively huge and draggy footprint in the cell, mitochondria are somehow nimbly towed about by one or two minuscule motor protein tethers through a dense tangle of the cytoskeleton.
It should not be that much of a stretch to go from the idea that the structure and behavior of mitochondria networks is an important part of cell division, to the idea they can control cell division, and therefore potentially control cancer in the differentiated tissues of multicellular organisms. Fission and fusion in mitochondria allow for cellular regeneration and avoidance of cancerous states. The retrograde response in mitochondria is critical when heteroplasmy rates rise so high that mitochondrial quality control is no longer controllable by normal pathway maintenance. Mitochondria appear to use two modes of energy and signal transmission: electrical/electrochemical and electromagnetic (optical). This signaling pathway is triggered by the loss of mitochondrial membrane potential. This means the loss of charge. This means too few electrons or too many protons or a combination of both. In talking to a noted medical geneticist and “single gene” champion Shane McKee, about the new review from Thomas Seyfried Ph.D. on the metabolic treatments for cancer, it is probably safe to say folks will be holding on to the somatic mutation theory of cancer for some time. Shane accepts that mitochondria play a significant role in cancer, but rejects the idea that they can be considered as ‘the root’ of cancer in the sense that the problem originates in mitochondria before it is evident in the genes. I suggest you reject Shane beliefs and reality in view of the 4 decades of Dr. Doug Wallace’s work on mitochondria.
People live their lives bound by what they accept as correct and true. That’s how they define their reality. But what does it mean to be “correct” or “true”? They are merely vague concepts. Their reality may all be a mirage for the public. Can we consider them to simply be living in their own world, shaped by their beliefs?
SUMMARY:
Melatonin regenerates the mitochondria respiratory proteins. Cancer is a state when the respiratory proteins are expanded and mitochondria have high heteroplasmy rates. Endogenous melatonin production form the eye and skin need the sun’s signal via these surfaces by way of neuropsin photoreceptor to regenerate our tissues using melatonin as a guardian protector of the respiratory chain in every human cell. The second messenger’s in this system in humans is the triplet state free radicals made by the respiratory proteins in mitochondria. The distance between each cytochrome and the amount of oxygen in mitochondria are quantized to determine what type of free radicals are made. These radicals can limit ELF-UV release from cells or increase it. Cancer is a state where ELF-UV light is released in massive amounts because we cannot retain the light because of lowered electric and magnetic fields being generated in our mitochondria. Ironically cancer cells cannot live without brisk ECT function. ECT speeds and endogenous ELF-UV light release are linked. When this occurs AMPk is also activated by change in the anions of the Kreb cycle. It appears to be linked to fumerase defects. Nora Volkow also showed mobile radiation from cell phones increases AMPk signaling in cells.
My bet is to restore melatonin levels in the tissue to repair autophagy to get rid of the bad power plants. It is well-known UV light regenerates melatonin in the central retinal pathways. This is why AM sunlight improves sleep in all studies I have read.
Every morning you need the sun to charge separate water in your body in RBC’s and with melanin to create the battery that creates your life. Sunlight power becomes buried in the EZ of cells that surround every protein in your body. That life is created by a cadre of photochemical that do things we all rely on to liberate electrons and allow them to be programmed by sunlight to create your life. You need to expose all your surfaces to this signals to make sense of the world around you. The food and exercise guru’s that keep bathing in blue light and never see this message and blame foods and a lack of exercise for their ills. My Time #9 blog shows you what you might miss when you focus only on food and completely ignore light at night.

We must understand biology as it is today, and not confuse how it is with how we wish it to be. What is published in the textbooks about biology no longer applies? This perspective shift is counterintuitive for most. The universe can be deduced from the smallest part of the cell if one studied it closely enough. That is what I chose to do because it broadened my vision for medicine. The landscape looks different from every branch of the tree.
Make like the Sphinx every morning, while meditating in the morning sun, look to the East and ground and remain as connected to this environment as you can and food will never be your main driver. Light, water, and magnetism works to limit food influence at the confluence of your mitochondria and can help you build endogenous ocular and brain melatonin to treat cancer optimally. It’s that simple an Rx to regenerate your tissues by shrinking your respiratory proteins even from a cancerous state.

CITES:
- Light Shaping Life, Roeland van Wijk
- The Vital Question, Nick Lane
- https://www.youtube.com/watch?v=KwbIR2yUziw
- https://www.youtube.com/watch?v=ahlDLjf8c90
- http://www.biosciencetechnology.com/news/2014/07/total-darkness-during-night-key-success-breast-cancer-therapy?et_cid=4066343&et_rid=652401564&type=cta
- http://science.sciencemag.org/content/303/5660/1007
- http://m.medicalxpress.com/news/2015-07-mitochondria-oncogenesis-metabolic-reprogramming.html